
───先生の研究テーマである非対称分裂についてわかりやすく教えてください。
私たちの身体は、約30兆個もの細胞でできているといわれます。その莫大な数の細胞も、もとは1個の受精卵が分裂を繰り返してできたものです。分裂してできた細胞は、神経細胞、表皮細胞、血球細胞など約200種類以上にものぼります。
さて、細胞が分裂するとき、同じ種類の細胞だけをつくりだす(対称分裂)とすると、細胞の種類は増えないはずです。数多くの細胞ができるためには、ある細胞が分裂して生み出された2つの細胞(娘細胞)がそれぞれ異なる運命を持っていなければなりません。こうした分裂を「非対称分裂」と呼びます。
私たちは、この非対称分裂がどのようにして起きるのかを、線虫(C. elegans)を使って研究しているのです。
───線虫を使うのはなぜでしょう。
細胞がどう分化していくかを調べるには、どの細胞からどの細胞が分化したのか、その細胞系譜が明らかになっていなくてはなりませんが、高等生物では細胞の種類も数も多すぎます。その点、線虫は細胞の数が959個しかなく、1個の受精卵がどのようにして959個の細胞になるのか、その細胞系譜がすべてわかっています。しかも、身体が透明で、細胞が分裂する状態を顕微鏡のスライドグラスの上で観察することができます。さらに、受精卵から成虫になるまで3日間と短く、遺伝子改変も容易です。非対称分裂のメカニズムを知る上では好都合でした。その上、発生においてほぼすべての細胞が非対称分裂を行うことも、研究対象としては優れている点です。
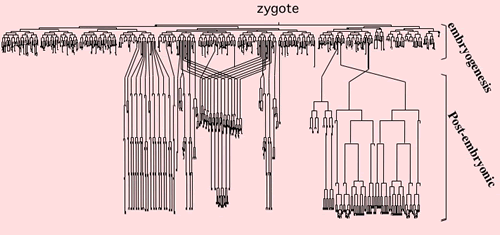
▲線虫C. elegansの全細胞系譜
───線虫の非対称分裂のメカニズムをどのように説き明かしていったのですか。
ホロビッツのラボメンバーのポール・スタンバーグが、一部の細胞で非対称分裂が起きず、同じ種類の細胞が2つできる変異体を発見していました。この異常の原因となる遺伝子こそが非対称分裂の謎を解くカギを握っているはずです。ラボの研究者仲間がその遺伝子の同定にチャレンジしましたが、成功していませんでした。そんななかで私は、LIN-17という受容体(タンパク質)の設計図となる遺伝子が変化したことが、この異常の原因であることを突き止めたのです。
その後研究を進め、線虫の非対称分裂においては、Wnt(ウイント)というタンパク質が細胞の表面のLIN-17受容体に結合して細胞内に伝わる「Wntシグナル」が重要な役割を果たしていること、そして、Wntシグナルを伝達するβカテニンというタンパク質が細胞内に偏って存在していることを、蛍光タンパク質で標識することよってビジュアルに示すことに成功しました。
この写真を見てください。明るい緑色に光っているのがβカテニンです。分裂時には細胞前方の表層に局在しているのに、分裂終期に新しい核が形成されるころには、後方の核へと蓄積していることがわかるでしょう。こうして、分裂してできた娘細胞が異なる細胞になるのです。
分裂初期
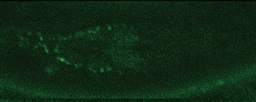
分裂初期
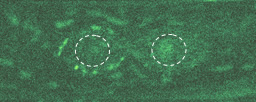
分裂初期
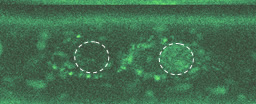
───なるほど、たしかに図の中の、核を示した丸の内部が明るく光っていますね。
線虫の多くの非対称分裂が、このWntシグナル経路で制御されています。興味深いことに、βカテニンのほか、がん抑制因子であるAPCも、同じようにWntシグナルによって細胞の前側の表層に偏っていくことがわかっています。
───いったいどのようなメカニズムで、βカテニンやAPCの局在が制御されているのですか。
詳しいことはまだわかっていないのですが、Wntシグナルに異常がある変異体では、βカテニンもAPCも局在が見られず、母細胞が分裂しても同じ娘細胞しかできません。
最近の研究で、細胞表層近くのβカテニンがβカテニン自身の核局在を抑制していること、そのプロセスに、APCと物質輸送を担う微小管が関わっていることを明らかにしましたが、そもそも細胞がWntのやってくる方向をどのようにして認識するのか、Wnt以外に方向を知るメカニズムが存在するかなど、まだまだ多くの謎が残っています。
▲Wntタンパク質が細胞表面の受容体に結合してWntシグナルを出すと、βカテニンとAPCが母細胞の頭側(図では左)に局在する。分裂終期から分裂後は、βカテニンは後ろ側の娘細胞の核(図右)に蓄積されるが、前側の娘細胞には蓄積しない。