ノックアウトマウスづくりに明け暮れる
———メモリアルスローンケタリングがんセンターで、ポスドクとして3年、その後研究員として2年研究を続けたあと日本に戻られます。きっかけは?
ずっとアメリカにいてもよかったんですけど、ボスがアリゾナに行くことになって。ちょっとアリゾナはな…と。もう一つ、日本に帰ろうと思ったきっかけになったのが、アメリカのセミナーで聞いたES細胞についての話です。ある大御所(当時は新進気鋭でしょうが)の先生のスピーチでES細胞からマウスがつくれるという話を聞いて、「そんなことができたらすごい、やってみたい」と思って紹介していただいたのが、今の筑波にある理化学研究所バイオリソースセンターの研究所です。井川洋二先生がつくられた研究所で、相沢慎一、八木健、古田泰秀、武田洋幸といったおもしろい人たちがいっぱいいました。そこでたしか、日本で初めて相沢さんがES細胞を使った実験を行ったんだと思いますが、ES細胞からノックアウトマウスをつくりましょうということになったのです。
———どんな遺伝子をノックアウトしたのですか?
テネイシンという遺伝子です。発生の初期から上皮と間充織(かんじゅうしき:胎生期の臓器が分化形成される過程であらわれる原始結合組織)の間にパーッと発現して、ものすごく重要そうに見えたんです。単なるノックアウトマウスではおもしろくないから、遺伝子の発現を可視化するためのマーカー遺伝子である大腸菌のLacZ(ラックジー)遺伝子を導入して、標的遺伝子の発現が見えるようにしようと工夫しました。
いや苦労しましたね。毎日シャーレ40枚の培地を交換して、細胞をスクリーニングして、ものすごい実験量でした。最終的には何とかノックアウトマウスができて私の役目は果たせたものの、残念なことに、明らかな表現型(フェノタイプ)が出なかった。遺伝子をひとつつぶすだけではだめだったんですね。それでまとめたのが、「ノックアウトマウスはできたけれど、何も起こりませんでした」っていう論文でした。
———その論文がノックアウトマウス作製の日本で最初の論文ですね。
はい。でも、ノックアウトマウスができたら、上皮と間充織の相互作用とか、どんどん研究が発展していくと思っていたのに、何も起きなかったので、それで研究は終りになってしまいました。もうショックで、特別研究員の任期も切れてしまって、いったいこれからどうしようというときに耳にしたのが、萬有製薬が新たな研究所をつくり、人を探しているという情報でした。それで何かのパーティの時でしたか、発生動物研究部長で、アメリカから帰国されたばかりの武藤(たけとう)誠先生にお会いして、「私、ノックアウトマウスつくれます!」って売り込んだのです。
———ノックアウトマウスづくりのプロとして、萬有製薬で活躍することになった。
はい。でも、理研で私が担当していたのは主に遺伝子をノックアウトしたES細胞を作る過程で、私自身はマウスをつくったことはなかったんです。一番難しいところはできるのだから、あとはたいしたことはないだろうと思っていたんですね。
ところが、遺伝子を導入したES細胞を初期胚にいれて母親マウスのおなかに着床させるんですが、おなかは大きくなるのに全然生まれないんですよ。やり方は合っているはずなのになぜだろうと、ずいぶん悩みました。原因は、単に移植する数が少なかったことでした。結局、マウスを育てる経験が足りなくてマウスのことを知らなかったわけ。
———その後は、とんとん拍子に?
ようやくマウスのシステムが完成して、次は何をノックアウトするかです。ES細胞の多能性はたしかにおもしろいけれど、やはり培養細胞でしょ。全能性をいうんだったら生殖細胞のほうがすごい。生殖細胞はもっとも重要な細胞であるにもかかわらず、その形成などはあまりわかっていません。「そうだ!生殖細胞をやろう」と、ターゲットが決まりました。

萬有製薬の仲間とエジプト旅行。1994年あたり?(左から2人目)
———企業の研究所って、そんなに自由に自分でテーマを決めることができるものなのですか?
そのころの萬有製薬って、ものすごく自由でした。当時の西村暹(すすむ)所長も「好きなことをやりなさい」と言ってくださって。無論、会社のプロジェクトもやっていましたよ。
そこで、生殖細胞に特異的な遺伝子を探していたときに見つけたのが、Mesp1という遺伝子でした。
———それで、Mesp1をノックアウトしたら、生殖細胞に異常のあるマウスが生まれたのですか?
いえ、そんなに単純じゃないんですよ。発生異常のフェノタイプは出ているんだけど、生殖細胞に関係はなかった。どこがおかしいのかと調べたら、どうも心臓がおかしいようなんです。そこでさらに調べていくうちに、Mesp1の隣に、Mesp2という遺伝子もあった。2つとも心臓形成に必須の因子だったのです。
そして、Mesp2は体節形成にも重要な役割を果たしていることがわかりました。ところが当初、Mesp2の発現を観察していると、何度やっても発現する検体と発現しない検体が出てくるのです。なので、実験は失敗だったなと思っていたんです。それが、よくよく探ってみると、失敗じゃなかった。
———失敗じゃなかったってことは、発現する場合と、発現しない場合と、両方とも正しいってことですか?
2時間のサイクルでオンとオフを繰り返していたんです! ヒトやマウスなどの脊椎動物には、脊椎骨とか肋骨などの繰り返し構造(分節構造)があります。発生過程で尾側から頭側へと波をうつように規則的に発現を変化させて細胞間の情報伝達が行われ、体節の分節する場所とタイミングを制御していたんですね。ダイナミックな生命現象をこの目で見ることができました。
野生型
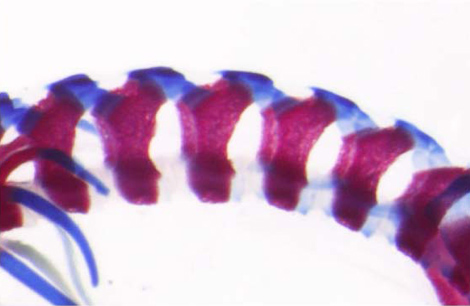
Mesp2ノックアウトマウス
Mesp2遺伝子をノックアウトすると、背骨や肋骨など分節構造に異常があらわれる。左が野生型、右がMesp2遺伝子のノックアウトマウス。分節化がおこらず、その結果脊椎骨が癒合している
———その後、萬有製薬から国立医薬品食品衛生研究所に移られますね。
企業に勤めたのは92年から97年までです。企業の研究所も良かったんですよ。だって、好きな研究ができたし、研究費の心配はしなくていいし、マウスの施設はすごく立派でしたから。でも、企業だとどうしても人事異動があって、長く勤めるうち研究職から離れざるを得なくなってしまうかもしれない。武藤先生も東京大学に移られましたし。研究環境としては快適でしたが、私もいいところがあったら移らなくちゃと思っていたときに、国立医薬品食品衛生研究所をご紹介いただいたのです。
しかもここでも何を研究してもよかった。毒性部という部署だったのですが、「毒にさらされると発生過程でいろいろな異常を起こすし、発生学を知らないと毒性学も深まらない」と言われて、ナルホドと納得しました(笑)。ラッキーなことに、萬有製薬で私がつくったマウスも全部持っていくことができたんです。ですから、衛生研究所でもノックアウトマウスづくりのテクニックをメンバーに教えて、いろいろなマウスをつくっていました。
2000年に今の国立遺伝学研究所に移ったのですが、遺伝研でもマウスとつきあい続けています。

衛研から遺伝研に移った直後(2000年)、体節形成の仲間(高橋淑子(右端)、武田洋幸、高橋雄ら)とフランスのオリビエ研を訪問