上皮細胞の接着構造から研究の道へ
———月田研究室に出入りするようになった経緯を教えてください。
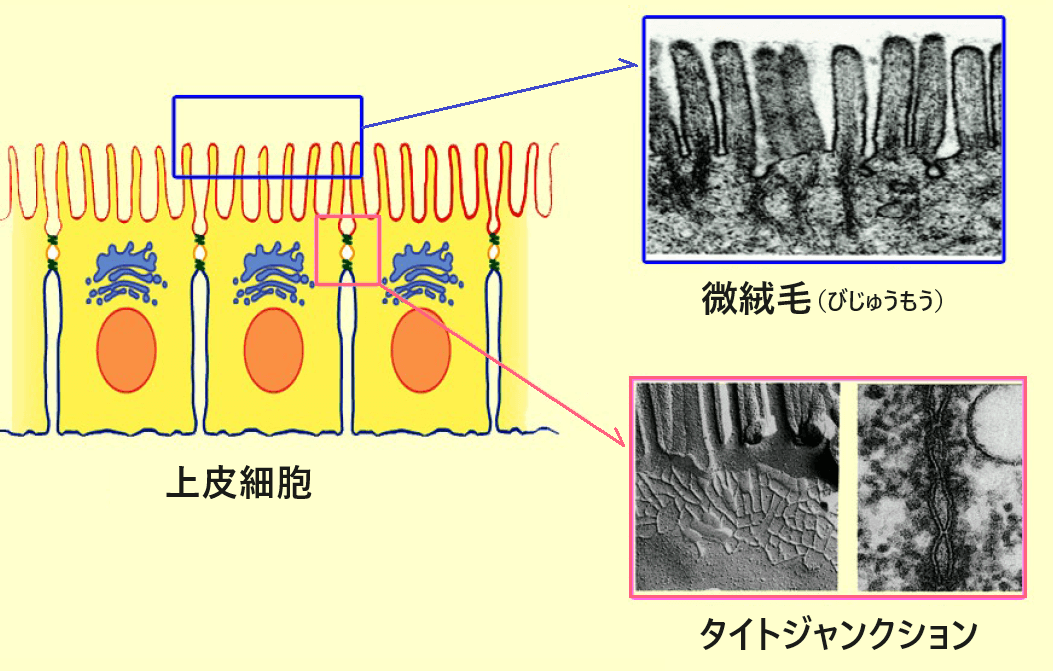
月田先生は上皮細胞の接着構造であるタイトジャンクションの第一人者で、電子顕微鏡を使って細胞の形態と機能を探る研究がご専門です。当時、タイトジャンクションを構成する主要なタンパク質の「クローディン」*を世界で初めて発見し、研究室は熱気であふれていました。ただ、私はそんなことは知らないまま、医化学の授業で「君たちは凡人なんだから、もっともっと勉強しなさい」と発破をかける先生の熱心な人柄に共感して、お話を伺うだけのつもりで研究室を訪ねていったのです。そうしたら成り行きで、研究室に通うことになりました。
*クローディン:タイトジャンクションを構成する主要なタンパク質の一つ。細胞膜上でクローディンが鎖状に連なり、隣接する細胞膜上に作られたもう一方の鎖とジッパーのように結合することによって、隣り合う細胞同士を密着させている。このクローディンの鎖が網目状になった構造体は「タイトジャンクションストランド」と呼ばれている。
上皮細胞とタイトジャンクション
上皮細胞は、私たちの皮膚や器官の表面にある細胞で、菌や異物が体内に侵入して感染症などを引き起こさないよう隣り合う細胞同士の隙間をピッタリ接着させている。タイトジャンクション(tight junction/密着結合)はそうした細胞接着構造の一つで、さまざまなタンパク質が結合し、複合体を形成している。
———研究室でどんなことをしたのですか。
まずは実験の手技を覚えなさいということで、当時、研究室にいらした永渕昭良(ながふち・あきら 現・奈良県立医科大学教授)先生のもとで、「カドヘリン」という細胞接着分子に結合するタンパク質の研究の手伝いをすることになりました。1年ほど経って、ようやく実験手技に習熟し、論文を読んで自分でも研究が進められるようになったころ、永渕先生は熊本大学に転出してしまいました。そこで月田先生にご相談して「研究室では多くの人がクローディンについて研究しているので、それ以外のテーマで考えたい」と申し上げたんです。すると「それなら、自由にやっていい」と。
———研究への道を踏み出したわけですね。
しかし、イバラの道です。無い知恵を絞っていろいろなアイデアを出したのですが、月田先生からはことごとくダメ出し。少し良いデータが取れて喜び勇んで報告に行っても「そのテーマは二番煎じ」「データの解析が甘い」など、厳しくも的を射た指摘に落ち込む日々が続きました。こんなことで仕事になるのだろうかといつも不安でしたね。しかしあるとき、上皮細胞が間葉細胞に転換する現象に着目して新しい接着分子を見つけたいと話したら、ようやく「やってみたら」とGOが出ました。
———間葉細胞とはどんな細胞ですか。
間葉細胞とは幹細胞のひとつで、iPS細胞やES細胞に比べれば能力は弱いのですが、骨、軟骨、脂肪などの細胞に分化することができます。胎児の発生期には上皮細胞が可動性の高い間葉細胞に転換して移動し分化を進めるのです。これを「上皮間葉転換」といって、通常、成人になるとその機能は眠ってしまいます。
———上皮間葉転換が胎児の発生期の分化を進めているんですね。
ところが2002年に、ある論文に出会いました。「スネイル」という遺伝子が上皮細胞に発現すると、カドヘリンの接着機能が抑制され、上皮間葉転換が起こって間葉細胞が自由に移動できるようになるというのです。
そこで、スネイルがカドヘリンの接着機能を抑制するなら、クローディンの接着機能も抑制するのではないかと考えて実験したところ、クローディンもスネイルによって抑制されることがわかり、論文にまとめることができました。ちなみに、スネイルは胎児期に間葉細胞が中胚葉や神経堤に分化するときに出てくる遺伝子で、これが何らかの理由で成人の上皮細胞に発現してしまうと、悪性腫瘍(がん)の細胞も自由に移動できるようになり、浸潤・転移につながることも報告されていました。
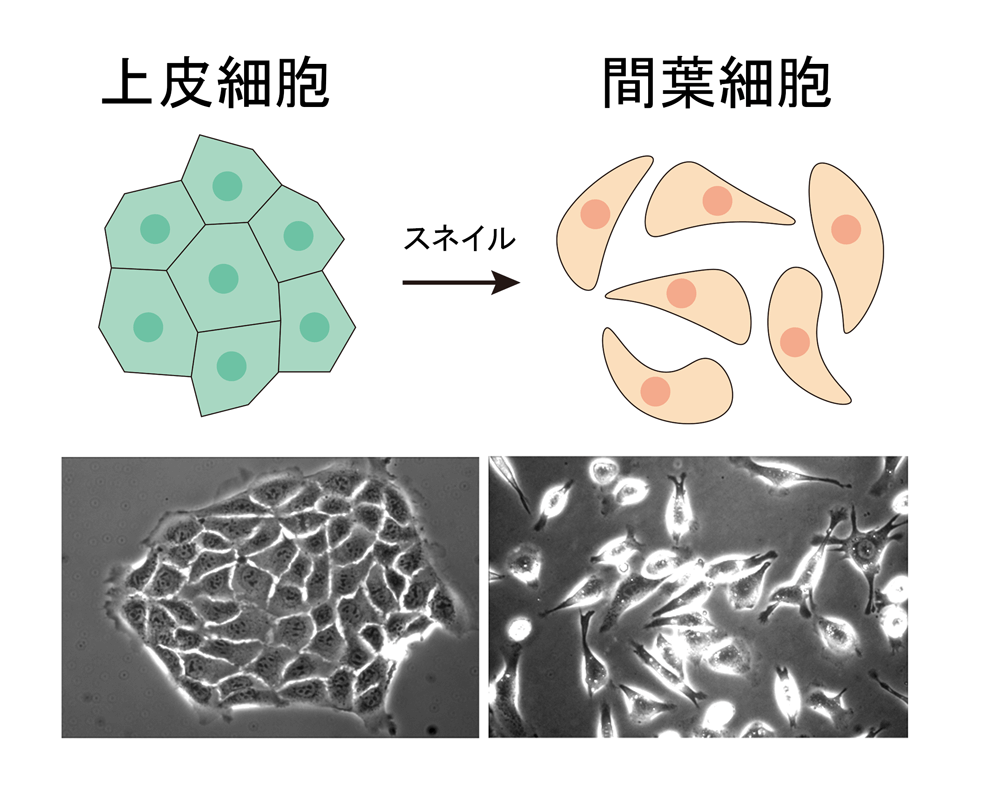
培養上皮細胞にスネイル遺伝子を強制的に発現させることによって、細胞同士が接着しなくなり、間葉細胞のようになる。